
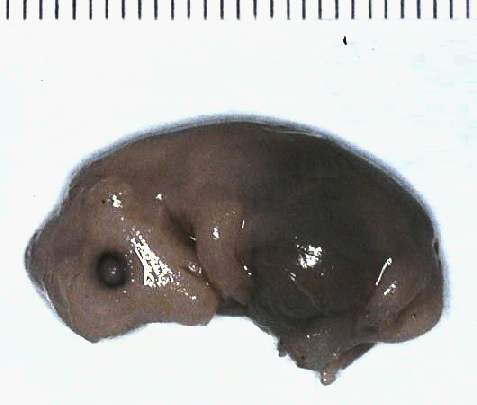
 Pig
Pig
The control that can be exercised over the prenatal growth of farm animals
is relatively slight, because the environment created for the fetus by
the uterus or cleidoic egg is buffered against changes in the mother's
environment.
A mammalian egg has virtually no yolk tissue (oligolecithal) whereas an avian egg contains a lot of yolk (telolecithal). Mammals evolved from reptiles with telolecithal eggs, and so they retain features that indicate that their early embryos once had to develop on a large inert yolk mass. The yolk mass of the poultry egg creates problems because it is thick and viscous, and cannot split into subunits as fast as the small cells of the developing embryo.
Cleavage is the process by which a zygote or fertilized
ovum subdivides into smaller cells called blastomeres. When a new
embryo has subdivided into a ball of eight or more blastomeres, but has
not yet formed layers of blastomeres, it is called a morula.
The morula starts to form a new animal from a clump cells called the inner call mass(A). Then the morula forms itself into a layer of cells called the trophoblast(B) surrounding a fluid filled space - the blastocoele(C) .
The trophoblast contributes to the placenta and is lost at birth. The
inner cell mass together with the trophoblast form a blasotocyst.
The embryo developing from the inner cell mass becomes roofed-over by amniotic
folds that later fuse to form a complete layer - the amnion.
Endoderm cells (F; which eventually form the gut and its associated
organs like the liver) spread from the inner cell mass over the inner surface
of the trophoblast and, at this stage, the blastocyst is said to be bilaminar
or two- layered.
The blastocyst becomes trilaminar or three-layered when mesoderm
cells (E; which eventually will form muscles, bones, and fat) migrate
from the inner cell mass spreads between the outer trophoblast layer and
the inner endoderm layer.
The mesoderm layer then splits internally, and a cavity expands within
the mesoderm to become the extra-embryonic coelom(G).
Expansion of the extra-embryonic coelom cavity allows the yolk sac and
allantois to expand into the coelom cavity from the gut of the embryo.
In poultry, the rate of growth of the extra-embryonic membranes may be
an important factor affecting overall growth so that, in genetic strains
of poultry with a high growth rate, the growth of the nutrient-obtaining
extra-embryonic membranes is faster than normal. Thus, paradoxically, although
overall embryonic growth may be slower in the strain with a high growth
rate, this allows growth to proceed at a faster rate than normal because
nutrients can be absorbed from the yolk at a faster rate.
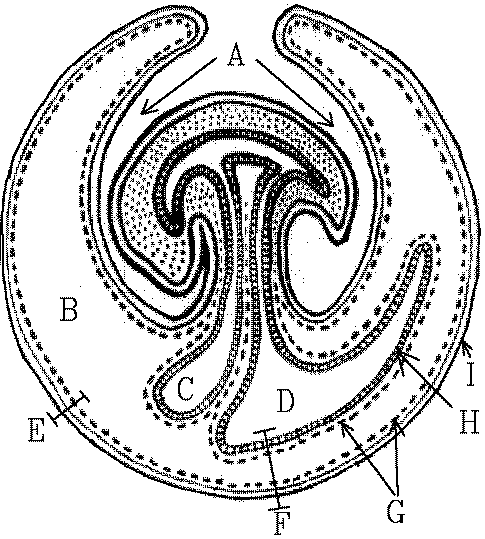
This is a horribly generalized view of how things fit inside each other:A = embry, B = coelom, C = yolk sac, D = allantois, E = two layers forming the chorion, F = four layers forming the allantochorion, G = two layers of mesoderm, H = endoderm, and I = ectodermal trophoblast.
In chicks, the yolk sac contains the yolk whereas in cattle,
sheep and pigs the yolk sac is rudimentary (because the developing mammal
gets most of its nutrients from the mother's uterus rather than from stored
yolk in a cleidoic egg).
In chicks, the allantois is used for respiration and storage
of waste products whereas in cattle, sheep and pigs, the allantois is developed
to provide vascular communication between the developing animal and its
mother.
The double layer of cells formed from mesodermal cells pushed up against
the ectodermal trophoblast is called the chorion. Likewise, the
allantoic membrane is pushed against the chorion to form the allantochorion.
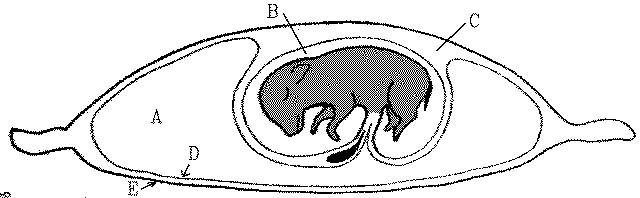
Membranes around a fetal piglet: A = allantoic cavity, B = amniotic cavity, C = coelom, D = allantois, and E = chorion.
The interface between the allantochorion and the mammalian uterus is increased in surface area by the formation of chorionic villi. In pigs, villi are scattered over the chorion to form a diffuse placenta whereas, in cattle and sheep, villi are grouped into about 100 separate mushroom-like structures called cotyledons.
The space within an avian egg is restricted so that the growth of the
fetus and allantois must be balanced against the shrinkage of the yolk
sac as its nutrients are used for the developing fetus. The mammalian uterus,
however, is capable of considerable expansion, which allows the allantoic
cavity to expand enormously.
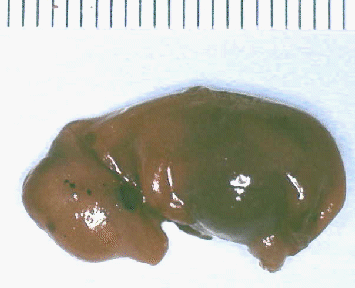 with this 45 to
50 mm embryo.
with this 45 to
50 mm embryo.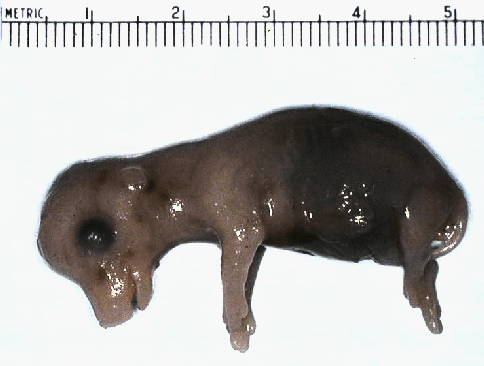 looking at the differences in relatives
sizes of the head, body, and limbs.
looking at the differences in relatives
sizes of the head, body, and limbs.
Now compare this 9-day chick 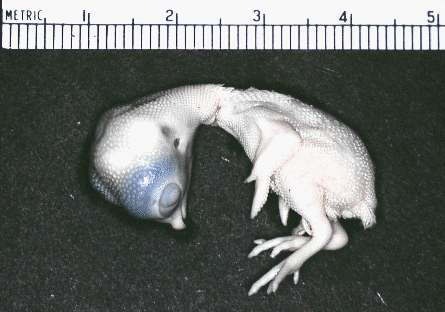 with this 12
day chick.
with this 12
day chick. using the scale to correct for differences
in magnification. See any feathers developing?
using the scale to correct for differences
in magnification. See any feathers developing?
Longitudinal growth of bones depends more on nutritional status and fetal body weight than on the age of the fetus. Maturation of the epiphyses, however, depends more on age than on nutritional status. The muscles of runt pigs are smaller than those of their more successful littermates. Runt pigs appear to have fewer muscle fibers, particularly white fibers, most of which are formed in the late fetal phase of fiber mass production. The smaller pigs of a litter, if they are able to compete successfully for milk after birth, may be able to catch up in their growth by the time of weaning. Below a birth weight of about 1 kg, however, runt piglets are doomed to produce carcasses with less muscle and extra fat.
Cleavage occurs as the zygote is passing along the oviduct before the egg is laid. The cytoplasm is concentrated in the blastodisc at the opposite pole to the yolk mass.
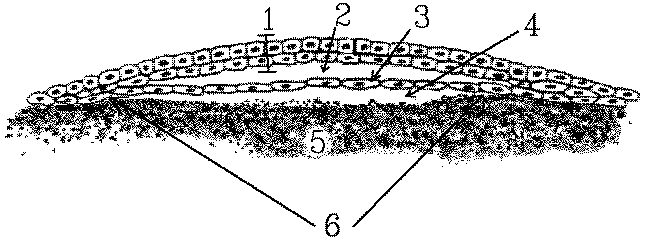
The blastodisc (sectioned in the diagram above)is a pale area several
millimeters across. A flat cavity called either the blastocoele
(2) or the cleft space develops within the depth of the blastodisc to produce
superficial (1 = epiblast) and deep (3 = hypoblast) layers
of cells. The presence of a subgerminal cavity gives the central part of
the blastodisc a semitransparent appearance so that this area is called
the area pellucida(6). The surrounding area (off the diagram) where
the edges of the blastodisc are in contact with the yolk is called the
area
opaca.
All the cells for the future chick embryo are recruited from the two layers of cells: the epiblast (1) and the hypoblast (3) in the area pellucida (6). The roles of cells from various regions of the epiblast and hypoblast in the developing embryo may be determined experimentally by marking the cells with nontoxic dyes.
Gastrulation is relatively simple in oligolecithal eggs, but in the flat blastodisc of poultry eggs gastrulation is more complex and the cellular intrusion characteristic of gastrulation occurs along a slit called the primitive groove . The cells (1) that will sink downwards and migrate sideways are located along a thick line of actively growing cells called the primitive streak(2) located between the epiblast (3) and the hypoblast (4).
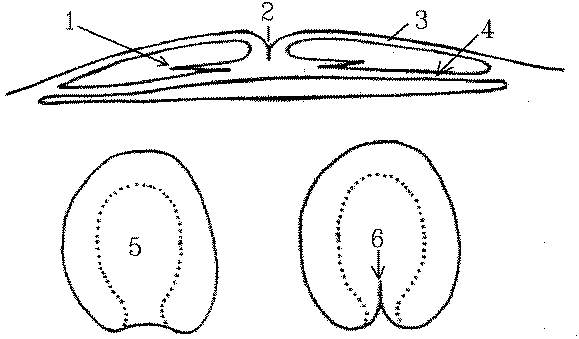
Top is a section, the two bottom diagrams are plan views showing
the area pellucida (5) starting to develop a primitive streak (6). At 24
hours, as shown below, the primitive streak is clearly visible.
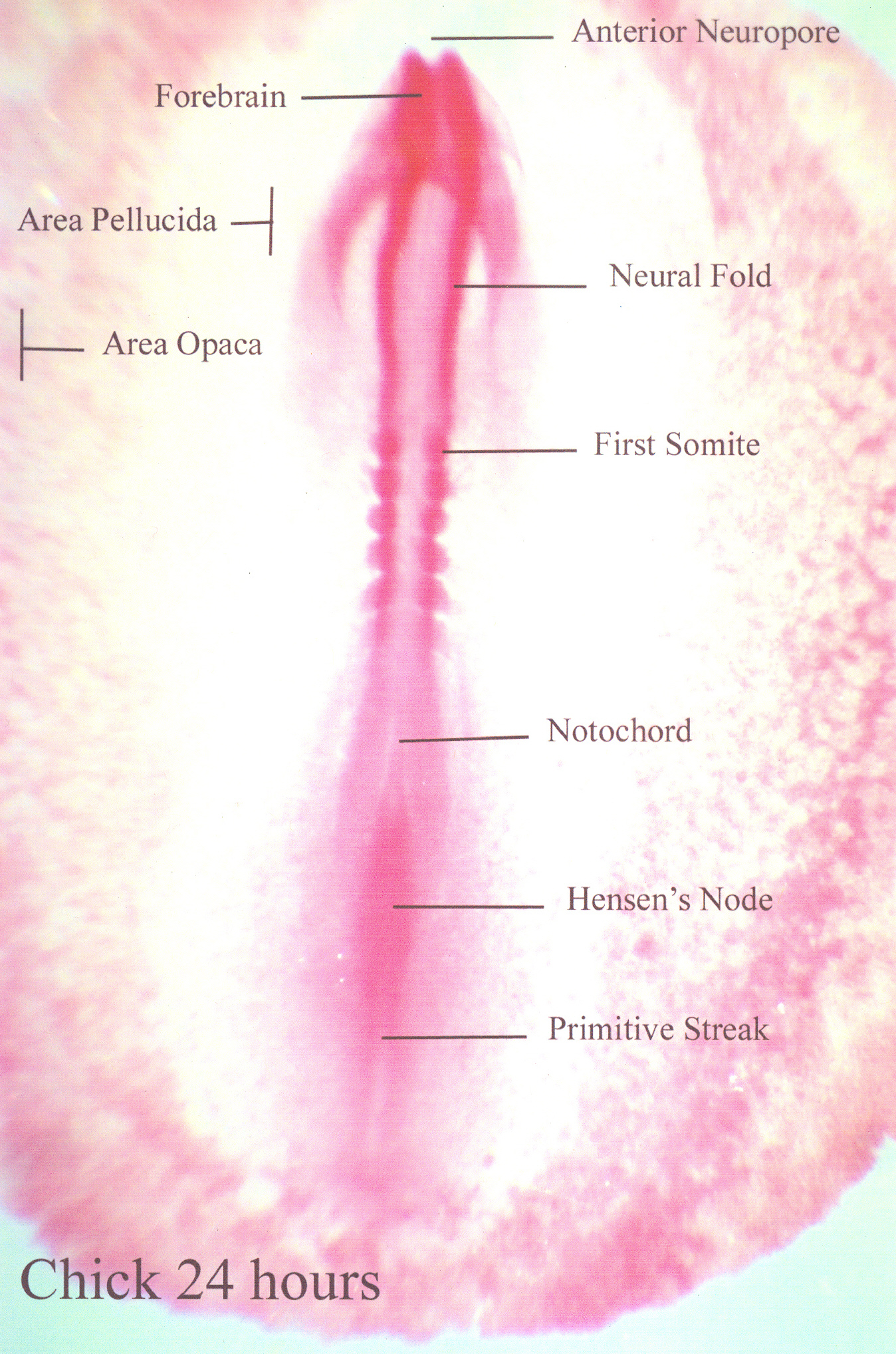
The primitive streak develops after the egg is laid and by about 16 hours of incubation is the most conspicuous feature of the developing embryo. The primitive streak indicates the location of the future vertebral axis. The mesoderm develops from a middle layer of cells that accumulates and spreads sideways below the primitive groove.
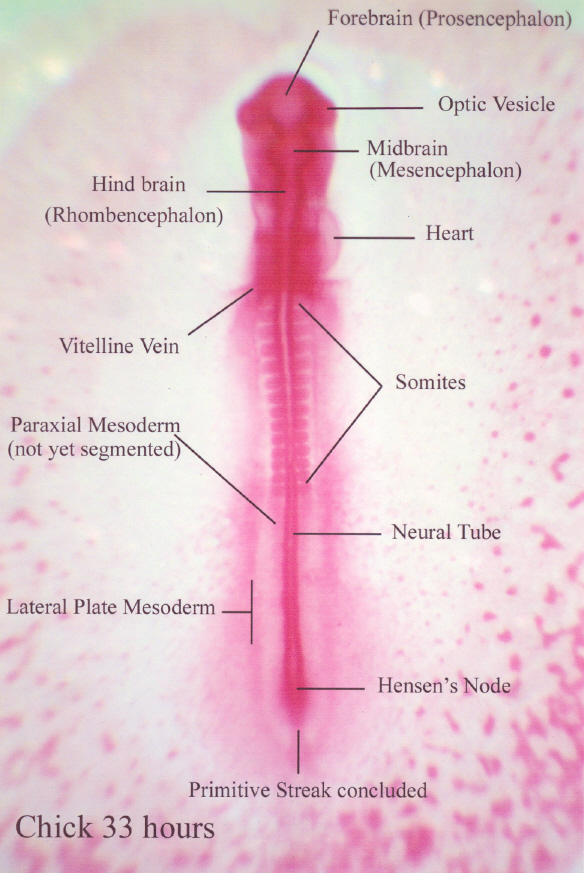
After the longitudinal axis of the chcik is established, primitive
streak activity is concluded. The embryo is lifted off the yolk by the
development of infoldings below the future head and tail regions of the
embryo. These infoldings unite along the sides of the embryo so that its
connection to the yolk is progressively restricted.

The amnion grows up and over, all around the embryo, and the amniotic folds then fuse to form a tent-like roof over the embryo.
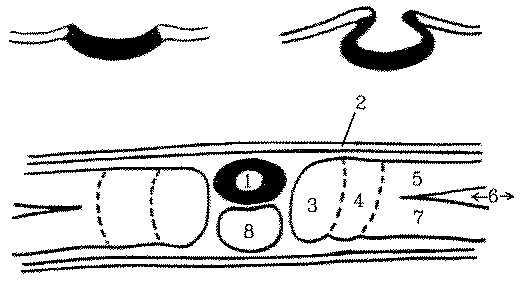
Sectioned chick embryos showing how the neural tube rolls up and sinks downwards: 1 = neural tube, 2 = epidermis, 3 = somite, 4 = intermediate mesoderm, 5 = somatic mesoderm, 6 = coelom, 7 = splanchnic mesoderm, and 8 = notochord.
The notochord establishes the future axis of the vertebral
column and is formed from some of the mesodermal cells in a middle layer
between the ectoderm and the endoderm along the primitive streak. Formation
of the notochord starts anteriorly and then posterior epiblast cells are
recruited as the notochord grows posteriorly.
Not all of the mesoderm within the primitive streak is used for notochord
formation and along each side of the notochord there remains a long strip
of mesoderm that develops into somitic and intermediate mesoderm. Laterally,
the remaining mesoderm splits into outer somatic mesoderm and inner splanchnic
mesoderm.
By this time, the nerve cord has developed from a sunken roll
of ectoderm sunken located above the notochord.
Understanding induction eventually may enable scientists to manipulate muscle development in the meat animals but, so far, attempts to identify the biochemicals responsible for embryonic induction have failed because of the infinitesimally small volumes involved. At present, we should be cautious in assuming that transmissible substances are solely responsible, because the messages that cause induction may be transmitted or faciliated by other means of cellular communication. However, the messengers of embryonic induction are most likely to be ribonucleoproteins.
Animal growth rates may be increased dramatically by introducing genes that lead to increased levels of growth hormone but whether or not genetic engineering at the molecular level will enable us to develop superior types of meat animals remains to be seen. The challenge is to make meat animals more efficient, not merely larger, and to enhance the quality of the meat. However, there is money to be made doing this so, probably, it will all happen and butchers one day will be cutting and selling meat cuts from designer animals (if we have not all been replaced by robots).
""