3 Embryology of Farm
Mammals
3.1 Duration of prenatal life
To keep things simple, we are going to call hatching -
birth. This will enable us to use prenatal to indicate events
occurring before birth in mammals and before hatching in poultry, and
postnatal for events after
birth or hatching. The duration of the
prenatal period is very important. Obviously, the producer needs
a good idea of when new animals will be born. A more subtle
point is the length of the prenatal period determines the cost of final
products. To produce a new calf takes a
certain length of time - a time during which all the costs of
production must be paid by the producer. Most of the animal's
live weight is obtained by postnatal growth. Thus, veal must be more
expensive per kilogram than beef - the prenatal period is the same for
both, but
beef gets a magnifier effect from longer postnatal growth.
Lamb must be more expensive per kilogram than mutton.
| Stage of development |
Beef
|
Lamb
|
Pork
|
Chicken
|
Forelimb bud
|
24
|
20
|
16-17
|
2.2
|
Hind limb bud
|
26
|
21
|
17-18
|
2.2
|
Foetus
|
45
|
21
|
20
|
5
|
Birth
|
285
|
150
|
112
|
22
|
Slaughter
|
850
|
350
|
270
|
70
|
Prenatal duration %
|
33
|
43
|
42
|
31
|
- The data in the table are days approximately. Gestation
varies between breeds and slaughter animals vary in age, depending on
market conditions. But
the percentages in the bottom line are close to commercial
approximations.
- Our control over prenatal growth is minimal because the
environment created for the foetus by the
uterus or cleidoic egg is
buffered
against the farm
environment.
- A
cleidoic egg is one that, except for temperature and the exchange of
gases, is
isolated from its environment by membranes and a shell, as in poultry.
3.2 Embryology
- Oligolecithal versus telolecithal eggs.
Mammalian eggs are oligolecithal - they
contain very little yolk. Poultry eggs are telolecithal -
they contain lots of yolk. In
a telolecithal egg, cell division in the relatively small amount of
protoplasm
does not spread very far into the viscous yolk mass. The future chick
develops as a flat disk of cells embedded on the surface of undivided
yolk. Mammals have very little yolk but have
evolved
from reptiles with telolecithal eggs and, thus, they retain
features to cope with the problem of a large
inert
yolk mass.
- Cleavage.
Cleavage is the
process by which a zygote
(fertilized ovum) is subdivided into smaller cells called blastomeres.
- Morula. When a
new embryo has subdivided into a ball of eight or more blastomeres, but
has not
yet formed layers of blastomeres, it is called a morula.

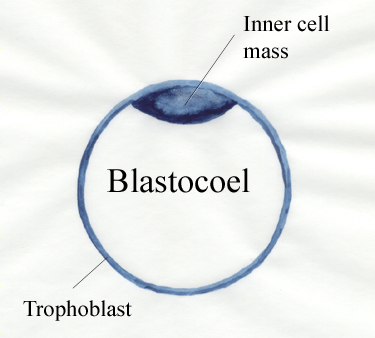
- Inner cell mass versus trophoblast. The morula
starts to
form a new animal from the inner cell
mass in the morula. The morula also
forms a
fluid‑filled sphere of cells called the trophoblast. The fluid-filled
cavity is called the blastocoel.
The trophoblast
contributes to the placenta and is lost at birth. The inner cell mass
together
with the trophoblast is called a blasotocyst.
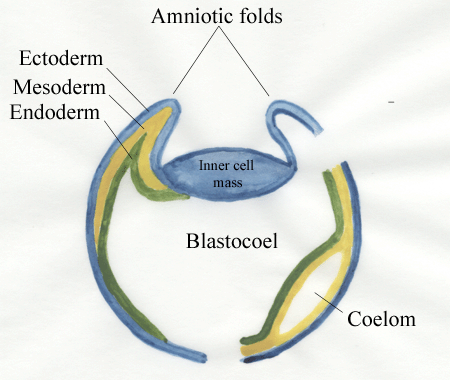
The embryo will develop
from the
inner cell mass roofed‑over by amniotic folds - these later fuse.
Endoderm from
the inner cell mass spreads over the inner surface of the trophoblast
and, at
this stage, the blastocyst is said to be bilaminar. The blastocyst
becomes
trilaminar when mesoderm from
the inner cell mass spreads between the
outer
trophoblast layer and the inner endoderm layer. The
mesoderm layer
then splits within itself to form a cavity within the mesoderm, the
extra-embryonic coelom.
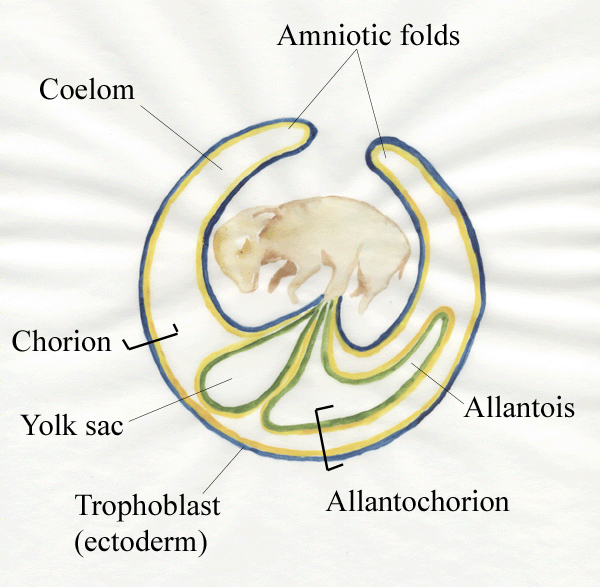
- Expansion of the
extra‑embryonic coelom
allows the yolk sac and allantois to expand into the coelom
from the
gut of the
embryo. The rate of growth of the
extra‑embryonic
membranes may be an important factor affecting overall growth. Thus, in
animals with a high growth rate, growth of the
nutrient‑obtaining
extra‑embryonic membranes is faster than normal. Initially, embryonic
growth is
slow in animals with a high growth rate, but later it proceeds
faster than
normal as nutrients are assimilated at a faster rate.
- In chicks, the yolk sac contains the yolk whereas
in
cattle, sheep and
pigs the
yolk sac is rudimentary.
- In chicks, the allantois is used for
respiration and
storage of waste products whereas in cattle, sheep and pigs, the
allantois
enables vascular communication between the developing animal and its
mother.
- The double layer of cells formed by the mesoderm and the
ectodermal
trophoblast
is called the chorion.
- The allantoic membrane is pushed against the
chorion to
form the allantochorion.
- The interface between the
allantochorion
and the mammalian uterus is increased in surface area by the formation
of chorionic villi.
- In pigs, villi are scattered over the chorion
(diffuse
placenta) whereas, in cattle and sheep, villi are grouped into
about
100
patches called cotyledons.
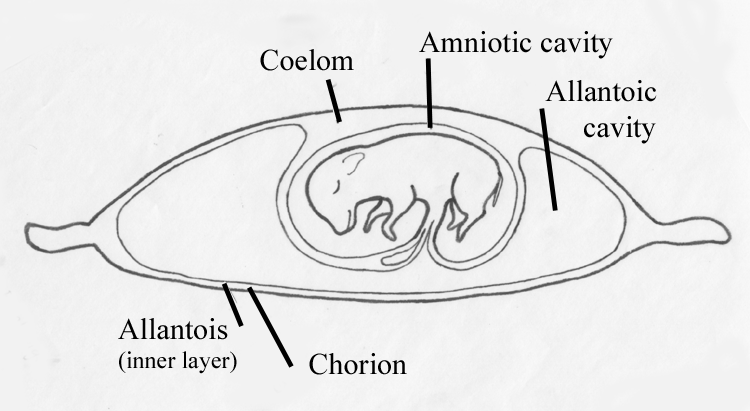
The space within an
avian egg is limited,
and the growth of the foetus and allantois is balanced by shrinkage of
the yolk
sac. The mammalian uterus, however, is capable of considerable
expansion, and
the allantoic cavity grows quite large.
3.3 Cattle
- The most widely used
measure of embryonic
and ofetal growth is crown-rump (CR)
length, a straight
line
measured from the crown of the head to the base of the tail. The crown
may be
taken as a point midway between the eye orbits on a line traversing the
approximate position of the frontal eminence.
- Growth in weight
of
the calf is usually more
variable than
CR length. The weight of the
foetus does not usually overtake the weight
of the
foetal membranes until approximately three months after conception.
- The
relative water
content of muscles decreases during foetal development.
- The
concentration of DNA
and RNA remains high until approximately 6 to 7 months gestation: after
this
time, the relative DNA content sharply declines and there is an
increase in dry
matter and total nitrogen.
3.4 Sheep
- In foetuses of unknown
age, CR length may be
used to determine the age of the foetus from 50 to 100 days
gestation.
- The age
of
foetal lambs also may be estimated from the state of development of the
hair
follicles in the skin.
- Undernutrition of the ewe during
the last
six weeks of gestation causes a severe decrease in the birth
weight of
lambs.
- Embryonic growth and
development
proceed in an anterior to posterior sequence with the head reaching a
relatively large size early in development. During foetal development,
the
remainder of the body catches up to, and overtakes the growth of the
head. The
head to body ratio, however, is rather variable between animals.
- The
feet and
the tail develop last.
- Birth weight has a
strong effect on the
early postnatal growth rate of lambs which slowly diminishes with age.
- Competition between
twin and
triplet foetuses may inhibit growth. In competitive situations, males
sustain less
inhibition of
growth than females.
3.5 Pigs
- Sows are
multiparous and
usually have ten or more offspring in each litter.
- Wild pigs have a
longer
gestation period (130 days) and fewer offspring per litter than modern
commercial pigs, so selection for larger litters
may have
decreased the length of gestation.
- Breeds with
few
piglets per litter have embryos with low embryonic growth rate and
fewer cells.
- Foetuses compete with each other for the
nutrients made
available by the uterus, and the success of a foetus is related to its
position
within the uterus.
- Selection for muscle
growth in pigs may have resulted in piglets being heavier but less
mature at
birth.
- Longitudinal growth
of bones depends more
on nutritional status and foetal body
weight than on the age of the foetus.
- Runt
pigs (the smallest in a litter) have fewer myofibres than normal.
- If runts are able
to
compete successfully for milk after birth, may be able to catch up in
their
growth by the time of weaning.
- But, below a birth weight of about 1 kg, runt piglets are doomed
to produce carcasses with less muscle and extra
fat than normal.
- Splayleg is a condition
in which newborn piglets cannot bring their hindlimbs together
normally. Thus, their locomotion is impaired and they may be crushed by
the sow.
3.6 Genetic engineering
The experimental
manipulation
of prenatal development in farm animals has resulted in commercial
methods such
as embryo transfer. This early
stage of animal
development is also the most amenable stage for genetic engineering.
Genes were
first
transferred experimentally into new locations by techniques such as
chromosome
assimilation and cell fusion.
To show the transfer of genes
actually had
taken place, the recipient cell was placed in an environment in which
the
transferred genes would reveal themselves. For example, the recipient
cells
might suddenly acquire the ability to survive in a medium that lacked a
hitherto essential nutrient. Other techniques also were developed for
the
transfer of isolated DNA: (1) direct
injection into the nucleus of a
recipient
cell using a micropipette or high-speed impact, (2) the use of a virus
to carry
the DNA into the recipient cell, and (3) precipitation of the DNA with
calcium
phosphate to make it attractive for uptake by the recipient cell.
The genetic
engineering of
mammalian development by such techniques has only just started and, at
the
present time, depends heavily on the use of relatively simple and
extensively studied genes. Apart from important exceptions such as
myofibre
hyperplasia in double-muscled cattle, most of the commercially
important
traits of
meat animals
are regulated by large numbers of genes and are difficult to manipulate
by
genetic engineering. In laboratory animals, growth rates may be
increased
dramatically by introducing genes that lead to increased levels of
somatotropic
hormone (STH). Whether or not genetic
engineering
at the molecular level will enable us to develop superior types of meat
animals
remains to be seen. The challenge
is to make meat animals more
efficient, not
merely larger, and to enhance the quality of their meat - not to scare
the
meat-eating public towards vegetarianism.
Further information
Structure and Development of Meat
Animals and Poultry, Chapter 8.