18 Adipose Tissue
18.1 Introduction
In the overall balance sheet for energy in agriculture, relatively
large amounts of feed energy are used when animals deposit fat in their
bodies, but much of this fat is removed and wasted after the animal is
slaughtered. About 6% of the live weight of a steer may be removed as
fat in the abattoir, and an equal amount may be trimmed from the
dressed carcass by the butcher.
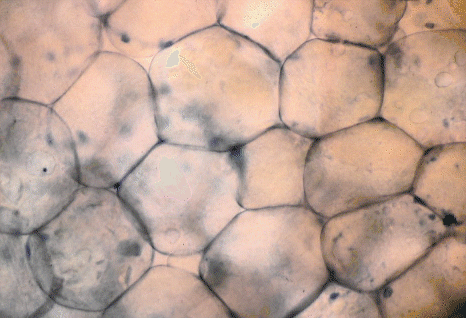 Adipose tissue, the cells
of which are shown above, only serves its proper function when an
animal uses the energy and insulation provided by its adipose tissue to
survive a period of inadequate feed intake or cold weather. Adipose
tissue in meat, however, is not altogether undesirable and wasteful. It
is desirable in moderation to give a "finished" appearance to a carcass
and, without at least some subcutaneous fat, a carcass is judged to be
unattractive by traditional standards.
Adipose tissue, the cells
of which are shown above, only serves its proper function when an
animal uses the energy and insulation provided by its adipose tissue to
survive a period of inadequate feed intake or cold weather. Adipose
tissue in meat, however, is not altogether undesirable and wasteful. It
is desirable in moderation to give a "finished" appearance to a carcass
and, without at least some subcutaneous fat, a carcass is judged to be
unattractive by traditional standards.
Fat deposited within
muscles (intramuscular adipose tissue) appears as
a delicate pattern of wavy lines in the meat - hence its common name, marbling fat. It is traditionally
thought marbling fat contributes to the juiciness of cooked meat
because it melts away from between bundles of myofibres to make the
meat more tender and more succulent. However, even though I agree with
this intuitively, it is difficult to find much scientific evidence in
support of this traditional view, except in poultry where subcutaneous
and intermuscular fat baste the meat as it is being roasted. Steaks
without marbling may have desirable palatability after rapid, high
temperature cooking, but only if they have been properly conditioned or aged (21 days). Unscientific as it
may be, there is a very sensible reluctance on the part of many
butchers to allow completely lean beef onto the market as top quality
meat, unless there is some redeeming factor such as extreme youth
(heavy baby beef) or excellent conditioning (21 days or more).
Much of the characteristic flavour associated with different
types of meat originates from carbonyl compounds concentrated in the
adipose tissue. But the flavour of meat also may be modified by the
animals' diet. Not all the fat in the carcass is macroscopically visible
(visible by eye without a microscope) because many myofibres,
particularly those in postural muscles, contain large numbers of
microscopic fat droplets. The storage of fat droplets within myofibres
is related to the overall metabolism of the animal. For example, the
deposition of lipid in the postural muscles of cows reaches a peak at
about one week after calving when lipid stores in the liver also are at
their highest.
The ancestors of present-day farm animals once were used to feed
populations of people, many of whom were manual workers who expended
large amounts of energy in tasks that we now perform by machine. Fat
contains more stored energy than lean, and the high fat content of meat
once supplied much of the energy that people expended in their daily
work. Now this extra energy is undesirable, and a reduction of the
fat content of meat is a major goal in the continued improvement of
meat animals.
Most of the meat eaten in the 19th century was derived from older
animals than are marketed now, and most of these animals had already
given long service in the production of milk or wool, or had been used
to pull ploughs or wagons. The presence of marbling fat would have
greatly improved the palatability of the tough and strong tasting meat
derived from these mature animals. Also, large families were more
common a hundred years ago, and large joints of meat cut from large
carcasses were well suited to domestic requirements.
To understand how meat may be made leaner, we need to look at the
origin, metabolism and proliferation of adipose cells.
18.2 Adipose cells
- A mature adipose cell or adipocyte may have a
diameter of about 0.1 mm and is filled with triglyceride. Thus, its nucleus and
cytoplasm are restricted to a thin layer under the cell membrane.
- When isolated from their surroundings, adipose cells are
rounded in shape but, when packed together in adipose depots, they are
compressed with flattened sides.
- Pockets of very small adipose cells sometimes appear between
normal-sized cells and this may bias measurements of mean cell size,
depending on how well the small cells are detected.
- Mature adipose cells have very little cytoplasm, which is why
the water content of fat is so low. Think about this for a minute. When
an animal makes 20 grams of meat protein, it adds them to 80 grams of
water to make meat. But when it makes 20 grams of triglyceride, it can
only add 1 gram of water to make fat. So this is why fat
is an economic disaster: triglyceride not only has a high energy
content but, even worse, it carries no water for free!
- In
many locations in the body, large numbers of adipose cells are grouped
together to form adipose depots.
- Adipose cells are kept in place by a meshwork of fine reticular
fibres (Type III collagen fibres).
- Large adipose depots usually are subdivided into layers or
lobules by partitions or septa
(singular is septum) of fibrous connective tissue.
- In the layered subcutaneous fat of a pork carcass, the septa
may follow the body contours and may create a weak boundary layer echo
in the ultrasonic estimation of fat depth.
- Adipose depots are well supplied by blood capillaries, which
are readily visible if the animal is not properly bled out at
slaughter.
18.3 Origin of adipose cells
- Speculation concerning the origin of adipose cells has continued
since the end of the 19th century, by which time most of the feasible
possibilities had already been proposed. In certain locations in the
body of a foetus, where adipose tissue will develop later in life, mesenchyme
cells congregate in lobules resembling glandular tissue (in an embryo,
the mesenchyme is a diffuse tissue composed of cells that may
differentiate to form the connective tissues of the body, as well as
blood and lymphatic vessels).
- The mesenchyme cells in gland-like
lobules begin to accumulate small droplets of triglyceride. The
droplets coalesce as they are crowded together and, finally, they form
a single large mass in the center of each cell, thus a multilocular (many lipid droplets)
structure has been replaced by a unilocular
one (one large lipid droplet). The cells are called gland-like
because they have many features typical of gland cells but are not
producing glandular secretions.
- In other locations where adipose tissue is about to develop, the
adipose cell precursors (sometimes called preadipocytes) may resemble fibroblasts (the cells
producing connective tissue fibres). In this case, cells are
spindle shaped with an oval nucleus. We must call them fibroblast-like
cells because we cannot prove they form connective tissue fibres like
true fibroblasts.
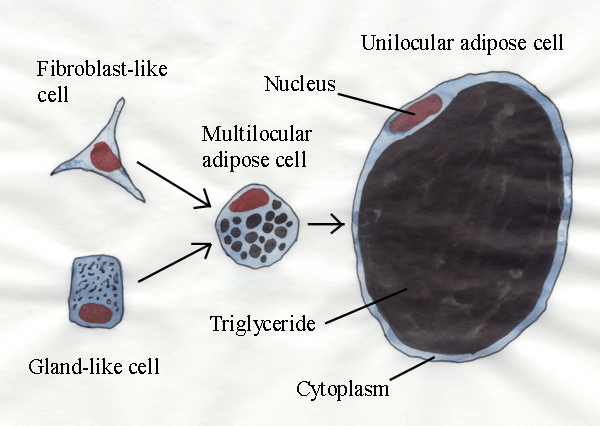
It is difficult to prove the gland-like
and fibroblast-like cells
giving rise to adipose cells are distinct types of cells with a rigid
pattern of development. However, the possibility cells are rigidly
programmed to develop exclusively into adipose cells is supported by
the behaviour of transplanted cells. Cells
transplanted from precursor adipose lobules to future low-fat regions
will store triglyceride and become adipose cells, even in an
inappropriate location. Similarly, precursor adipose cells from
certain sites will continue their programmed development into adipose
cells, even if they are removed from the body and cultured in the
laboratory. However, the trigger to differentiation appears to be
hormonal because it is influenced by insulin,
thyroid hormones and
insulin-like growth factor (which is called IGF-1 and is a hot topic for
research right now). The trigger or inducer acts upon a committing gene
for the differentiation pathway.
One source of adipose cells may be the recruitment of fibroblasts.
Fibroblasts can increase in number by mitosis. When recruited to become
adipose cells, they give up the shape and activities of a fibroblast,
and pass through a multilocular stage of triglyceride accumulation,
finally to become adipose cells resembling those derived from precursor
cells. Adipose cells sometimes originate from the endothelial cells of the blood
vascular system and there is evidence of this in pigs. Although extra
adipose cells may be recruited when animals become obese, the extra
cells are retained when an animal returns to its normal level of
fatness.
18.4 Adipose tissue distribution
Adipose depots range in size from small groups of adipose cells
located between muscle fasciculi (bundles of myofibres), to the vast
numbers of adipose cells located subcutaneously
and viscerally. It is
important to distinguish between anatomical
sites and systemic
locations. Specific muscles or regions of the carcass are
anatomical
sites. The following are systemic locations.
-
Intermuscular (between muscles).
-
Intramuscular (within muscles).
-
Visceral (around the guts).
-
Subcutaneous (under the skin).
For example, fat from a specified anatomical site such as the shoulder
may be separated into different systemic depots (subcutaneous,
intermuscular and intramuscular). The distinction between anatomical
sites and systemic locations is important commercially. For example,
bovine intramuscular marbling fat sometimes first becomes noticeable in
rump and loin muscles, where it adds to their value. In the same
systemic location, but at a different anatomical site, such as the
brisket (cut of meat including the sternum), marbling fat confers no
economic advantage and is wasteful. In cattle, the relative growth of
subcutaneous fat is similar in both the forequarter and the
hindquarter. However, the relative growth of intermuscular fat is higher
in the forequarter than in the hindquarter. There is no guarantee
systemic fat depots are homogeneous, even at a single anatomical site.
For example, the backfat seen on pork carcasses is subdivided into
three layers differing in their composition and pattern of growth.
The systemic deposition of fat in a carcass influences commercial
indices of carcass composition such as the dressing percentage. Intramuscular fat present
in meat at the time of cooking is mostly retained within the meat. The
total of omental fat,
mesenteric fat, and kidney fat may constitute
about 30% of the total fat
in a beef steer.
- The omentum is a thin
connective tissue sack filled with fat and located over the intestines.
The omental fat is often called caul
fat.
- The mesenteries are thin
connective tissue membranes supporting the intestines.
- Kidney (renal) fat is a
knob of hard fat around the kidneys - it may be sold as suet fat.
DRESSING % =
(CARCASS WEIGHT / LIVE WEIGHT) %
Thus, most of the fat deposited around the viscera is removed with
the viscera at slaughter, and this reduces the dressing percentage. Fat
deposited between or within carcass muscles increases the dressing
percentage.
In cattle, it was traditionally maintained thought fat deposition
followed three systemic phases.
- In the first phase, fat was thought to be deposited around the
viscera and kidneys, and within the caul and mesenteries.
- In the second phase, fat was thought to be deposited
subcutaneously and intermuscularly.
- The third phase was thought to be the deposition of marbling
fat within muscles.
But, with modern cattle, no simple chronological separation of the
three phases of fat deposition is detectable, and the relative amounts
of fat in the main systemic locations may remain constant. On a high
energy ration, cattle may deposit subcutaneous fat at a greater rate
than they deposit intermuscular fat. However, this difference does not
appear when animals are on a low energy ration. Breeds of cattle differ
in the way they develop their systemic fat depots. For example,
Herefords may produce more subcutaneous fat and less perirenal and
pelvic fat than Angus, Friesian and Charolais crossbred cattle.
However, when adipose growth at different anatomical sites is examined,
relative to total fat, only minor differences generally are found
between breeds.
In pigs, subcutaneous fat grows at the same rate as total body fat,
intermuscular fat grows more slowly, and visceral fat grows faster. The
separation of phases in adipose deposition is complicated. While the
experimenter usually works in terms of calendar days and weeks, the
experimental animal is following its own physiological calendar. The animal's
physiological calendar is based on events that mark the progress
through its life cycle. For example, skeletal and reproductive
development follow an orderly sequence of events, and different breeds
may progress through this sequence at different rates. Measured in
calendar weeks and months, early-maturing breeds deposit noticeable
amounts of marbling fat before late-maturing breeds. Thus, the
introduction of late-maturing breeds with a large adult size may be
used to delay fat deposition and to enhance lean growth in cattle
populations.
Sex hormones may produce large and economically important
differences in the overall fatness of beef, mutton and pork carcasses.
Provided that comparisons are made at equal fatness, however,
- bulls, rams and boars generally produce leaner carcasses than
steers, wethers and barrows, respectively,
- steers and wethers generally produce leaner carcasses than cows
and ewes, respectively,
- in pigs the situation is reversed, and gilts generally produce
leaner carcasses than barrows, and
- the deposition of fat tends to occur at a lighter weight in
heifers than in steers, and at a lighter weight in steers than in bulls.
In general, triglycerides located subcutaneously where they may be
relatively cool in the live animal have a low melting point. Conversely,
perirenal or suet fat in the beef carcass is brittle at room
temperature since it comes from a warm place in the body and has a high melting point.
18.5 Reducing the fat content of meat
The genetic and nutritional factors regulating the amount and
distribution of adipose tissue can be described in terms of cellular hyperplasia (an increase in cells
numbers) and cellular hypertrophy
(an increase in cell size). An early stimulus to research on these
topics was provided by the hope of adipose cell numbers being
genetically regulated, while adipose cell size might be nutritionally
regulated. Had this been the case in meat animals, selection for
animals with low numbers of adipose cells might have provided a method
for enhancing the production of lean meat. It was also suggested that a
low plane of nutrition early in life might reduce the numbers of fat
cells, so animals would be less likely to deposit fat in the final
stages of growth to market weight.
Once adipose cells start
to store triglyceride, they are no longer capable of cell division,
and insulin causes increased adiposity by increasing the size rather
than the number of adipose cells. It has also been proposed adipose
cell size may be involved in the regulation of feed intake and
triglyceride deposition.
A basic cellular problem prevents reducing the number of
adipose cells by a low level of nutrition early in life.
Populations of outwardly identical adipose cells in adult animals are
formed from an initial population of specific precursor cells which is
variably supplemented by the recruitment of fibroblast-like cells.
Thus, the apparently limitless possibilities for recruitment of extra
adipose cells may allow compensatory growth to offset any reduction in
the initial population of specific precursor cells.
However, some possibilities do remain. In double-muscled cattle,
adipose cells are greatly reduced in number (cellular hypoplasia).
Since double-muscling is a genetic condition, there could be a simple
genetic mechanism regulating adipose cell numbers in these animals.
Also, adipose cell number and size can be modified by the selective
breeding of mice for either high or low postweaning growth rate.
Regulation of adipose cell numbers is affected by thyroid hormones but
not by growth hormone.
Passive immunization of meat
animals with antibodies developed against adipose cell membranes offers
a novel approach to the age-old problem of fat reduction in meat
animals. Perirenal fat and subcutaneous fat may be reduced in lambs by
this method without a detrimental effect on the efficiency of carcass
production, using three daily intraperitoneal injections. The technique
also works on pigs and has the added advantage of increasing muscle
growth.
The possibility of repartitioning
the flow of energy into parts of the body has been investigated with beta-agonists such as Clenbuterol. The fatty acids
released by beta-adrenergic stimulation of adipose tissue appear to be
diverted towards the provision of energy for protein synthesis. Thus,
muscle growth is enhanced and adipose tissue growth is reduced. The
effect may be enhanced by a simultaneous reduction in protein
degradation and an increase in growth horomone.
Another approach to reducing adipose accumulation is the suppression
of fatty acid synthesis by inhibition of gene expression, but how this
might affect adipose cell numbers and size remains to be seen.
18.6 Fat in pigs
- Newborn pigs have very little fat relative to other mammals.
- Between birth and 4 weeks of age, the percentage of fat increases
from about 1% up towards 18% of empty live weight.
- Adipose tissue lobules in regions that will later deposit large
amounts of fat contain few cells, relative to calves, and they are
separated by areas of loose mesenchyme or undifferentiated tissue.
- Triglyceride deposition in adipose cells first becomes
microscopically detectable at about the third month of gestation.
- After this time and for the remainder of gestation, the numbers
of cells in each lobule show no great increase, although the numbers of
lobules and the total numbers of adipose cells do increase.
- In pigs, fibroblast-like cells and gland-like cells are not the
only source of adipose cells, since perirenal adipose cells may
originate from endothelial cells of the vascular system. Fat is
deposited very rapidly around the kidneys, and perirenal fat has high
enzyme activity, large adipose cells and a low connective fibre content.
- The heritability of adipose deposition is quite high in pigs,
typically about 50% for back fat thickness, which explains the rapid
progress being made towards the production of very lean pork.
- Commercial selection for decreased backfat thickness produces a
real reduction in total carcass fatness, not simply a redistribution of
adipose tissue. However, the backfat of pigs is divided into two or
three layers (depending on the position of measurement) and the
different layers exhibit some degree of independence in their depth
changes in response to selection. The deepest layer may be most
responsive.
- Lean breeds of pigs may have a greater ability than fat breeds to
mobilize their fat.
18.7 Fat in cattle
- An increase in adipose cell numbers is mainly responsible
for
the postnatal accumulation carcass fat in cattle.
- As in pigs, visceral adipose tissue in cattle develops
before
intramuscular adipose tissue.
- Adipose precursor cells make their greatest contribution
to
cell numbers viscerally, while the recruitment of fibroblast-like cells
is more important intramuscularly.
- The increase in cell numbers (hyperplasia) is nearly
complete
in subcutaneous and perirenal depots by approximately 8 months. But,
within muscles, progressive cellular recruitment, may continue for much
longer, which is how the marbling of beef originates.
- Also, the balance between adipose cell hypertrophy and
hyperplasia might be influenced by animal nutrition. Steers fed hay may
produce less total adipose tissue with larger cells than steers fed a
high-energy ration. The animals on the high-energy ration may produce
more total adipose tissue, but with smaller cells. The difference
created by this nutritional treatment is simply one of a static adipose
cell population versus an actively growing population. Mean cell size
might be kept low by the formation of new cells in the active
population.
Marbling fat in the rib-eye muscle (longissimus dorsi) is important in
beef grading.
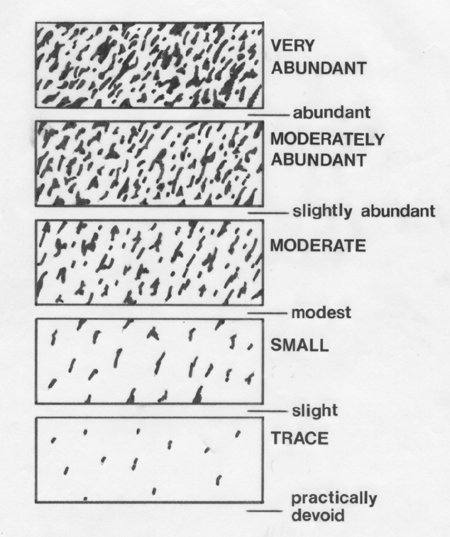
The
subjective assessment of marbling fat is very important in the
commercial evaluation of beef. Our basic problem is our customers
want to buy beef that looks very lean - even completely devoid of
marbling. But when we feed them steaks in a restaurant, they always
prefer the ones that were moderately or highly marbled. In other words
we have a real problem, because our customers have a taste preference
for beef that they don't like the look of!
18.8 Fat in lambs
Lambs have little prenatal development of subcutaneous fat, although
it develops rapidly immediately after birth.
18.9 Fat in poultry
- Avian adipose cells differ from those of farm mammals since they
have only a limited capacity
for lipogenesis - forming new triglyceride within the cell.
Thus, they rely mainly on the capture of circulating lipids synthesized
in the liver or released by digestion in the gut.
- There are marked differences in adipose cell numbers and
diameters between layer-type and broiler-type fowl, with the lean
broiler-type birds having fewer and smaller cells.
- An early restriction of nutrient intake, although it inhibits
adipose cell hypertrophy, has only a slight effect on adipose cell
hyperplasia. Thus, adequate cell numbers for fat deposition are present
in birds once they are returned to an ad libitum diet.
- Growth of the abdominal fat pad in chickens is from a combination
of adipose cell hyperplasia and hypertrophy, up to about 12 to 14
weeks, then it continues mainly by hypertrophy.
18.10 Steatosis
After cutting thousands of normal carcasses, it is always a surprise
to find one radically different in its fat distribution. The
accumulation of adipose cells is a common end result of a number of
pathological conditions affecting skeletal muscle. In most of these
conditions there is usually some evidence of myofibre regeneration. In
meat animals, however, it is not uncommon to find myofibres replaced by
adipose cells without any evidence of myofibre regeneration
and with no decrease in overall muscle volume. The most appropriate
name for this condition is muscular
steatosis. Muscular steatosis is most frequent in cattle and
pigs, but may also occur in sheep. Sometimes the occurrence of
muscular steatosis is indicated before slaughter by an abnormal gait,
but usually the condition is not found until a carcass is butchered.
The dividing line between excessive marbling fat and muscular steatosis
is sometimes difficult to establish, and it is often only the
restriction of muscular steatosis to a single muscle or muscle group in
an otherwise poorly marbled carcass making it conspicuous. Muscular
steatosis sometimes occurs in conditions which suggest it has
been caused by muscle damage or denervation. Strenuous muscle exertion
may cause extensive muscle damage, particularly in those muscles used
when an animal rears up on its hind legs. Steatosis can
sometimes be a major problem when it affects a high proportion of pigs
in a herd and, to the best of my knowledge, there is no scientific
explanation of the problem.
18.11 Conclusion
Fat is a lot more complicated than
it first looks, once we start looking at its cells under the
microscope. The big problem we have before us in the meat industry is
how to reduce the levels of fat in meat without leaving it dry and
tasteless when we eat it. Fat contributes a lot of taste to meat,
particularly flavours allowing us to recognize one species from
another. Without it, we may end up with just a bland, general meaty
taste.
Further information
Structure and Development of Meat
Animals and Poultry. Pages 107-131.