Bones account for a lot of waste, so they must be cleaned carefully and tediously to reduce economic losses, and butchers seldom look at them favourably. The utilization of bone waste is an important industry, but the give-away price of bones always seems too low to the butcher. The smell of hot-sawn bone or bone-taint from deep in a beef hip is not something to waft around the customers. Grudgingly, it might be admitted that bones give a shape to many traditional cuts of meat, and that traditional eating habits are something that many small butchers depend on for their livelihood. And we all know that meat close to a bone often has the best taste in a whole roast, and that bones help the cooking heat to spread evenly through a joint. But bones are a lot more important than this, if we take an overview of the whole agrifood system, from the farm to the dinner plate.
A meat animal's body is supported by bone, is held together by fibrous connective tissue, and is protected against starvation and cold by adipose tissue. These three types of tissue, although they differ radically in appearance and properties, are all classified as types of connective tissue. All three types contain cells located in a matrix with fibres.
In bones, both the matrix and the fibres make an important contribution to mechanical strength. The hardness of bone originates from a calcified matrix, and strength comes from embedded collagen fibres. The cells of bone, osteocytes, are trapped in small caves called lacunae. The gristle of the carcass is formed from tendons (by which muscles pull on bones), from ligaments (which hold bones together at the joints of the skeleton), from aponeuroses (which cover some muscles) and from fasciae (which form strong sheets between muscles). The dominant protein in gristle is collagen. Since connective tissues permeate nearly all parts of the body at the microscopic level, collagen is the most abundant protein in the animal body. The collagen fibres in meat are converted from strong fibres to jelly (gelatin) by the action of moist heat during cooking, but the collagen in bones may be removed by mild hydrolysis to produce gelatin for use in other food products or for other uses such as photographic emulsions.
In the section on skeletal anatomy, the number of vertebrae in pork carcasses was seen to vary between breeds. The heritability of the number of vertebrae is about 0.74 (which is quite high). Each extra vertebra adds about 15 mm to the length of the carcass at slaughter weight, so variation in vertebral numbers enables breeders to change carcass length. Breeds with a large size when mature and with heavy bone development tend to have more thoracic vertebrae than lighter breeds. Sometimes the ribs on extra thoracic vertebrae are only partially formed, but usually they are complete. The minimum number of lumbar vertebrae is generally found in cacasses with the maximum number of thoracic vertebrae. However, the variability of vertebral numbers frequently leads to an increase in the total number of vertebrae, so that the phenomenon is not due simply to the substitution of one type of vertebra for another.
But how is this variability created? Embryology tells us that the number of vertebrae in an animal is determined by the number of somites or tissue blocks that develops along the length of the spinal cord. By definition, in mammals, the vertebrae that bear ribs are identified as thoracic vertebrae. In the embryo there are ossification centers on each side of the developing vertebrae. In vertebrae that do not normally develop ribs, these lateral ossification centers contribute their bone tissue to the centra or bodies of adjacent vertebrae. In the thoracic vertebrae, however, thes laterally derived bone tissue remains separate from the centra and forms the ribs. Thus, the numbers of pairs of ribs and the numbers of thoracic vertebrae are determined by the developmental mechanism that controls the fate of the tissue which is derived from the lateral ossification centers.
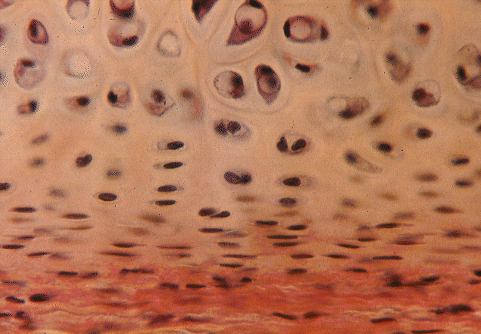
Cartilage cells, called chondrocytes, occupy lacunae in a stiff flexible matrix formed from collagen fibres embedded proteoglycan. Hyaline cartilage, with a white translucent appearance, occurs on the smooth surfaces of joints. In the larynx, trachea and bronchi of the plucks, hyaline cartilage forms the rings and tubes that hold these air ducts open during respiration. Flexible parts of the skeleton, such as the dorsal part of the scapula and the linkages between the sternum and the posterior ribs, also are formed from hyaline cartilage.
The importance of cartilage to the butcher is in helping to estimate the age of an animal at slaughter. Most of the bones of the carcass are initiated prenatally as cartilagenous models that subsequently become ossified. Complete ossification is a slow process, and the bones of young meat animals are more flexible than those of adults. Thus, the degree of ossification is a useful clue to animal age in the carcass.
Degree of ossification or its opposite, survival of cartilage, enables the carcass to be placed into a maturity group. This is done by examining a number of the features of the skeleton. In young cattle (around one year of age), the interiors of the vertebral centra are soft, red and porous in appearance. The medial surfaces of the ribs are rounded and streaked with red. As cattle grow older, the interiors of bones become harder, more white, and less porous. Carcasses from young animals exhibit a lot of relatively soft cartilage, particularly on the tips of the dorsal spines of the thoracic vertebrae. As animals grow older, cartilage in such locations becomes hard and ossified. In young animals, the sacral vertebrae are only loosely fused together, wheras older animals have their sacral vertebrae solidly fused together.
Chondrocytes are initially capable of both cell division (mitosis) and matrix formation. So clusters of related cells become pushed apart by their new matrix in a process called interstitial growth. Cartilaginous models of prenatal bones are covered by a membrane known as the perichondrium. Inner perichondrial cells differentiate into chondrocytes so that, in addition to interstitial growth, new cells and matrix may be added superficially in a process known as appositional growth. Cartilage may acquire numerous elastic or collagen fibres to become elastic cartilage or fibrocartilage, respectively. The dominant type of collagen in hyaline cartilage is what biochemists call Type II collagen and it accounts for 50 to 70% of the dry weight or collagen. But cartilage also contains some unusual minor types of collagen, such as Type M collagen, which is much shorter than other collagen molecules, and has a helical molecular structure that is stabilized by disulphide bonds.
Under the microscope, there are a number of features of cartilage that indicate an animal's physiological age.
(1) With age, the matrix becomes increasingly rigid.
(2) Appositional growth is the dominant growth process in older animals.
(3) Young chondrocytes are flattened or elliptical with their long axis parallel to the surface of the cartilage.
(4) Old chondrocytes are large (up to 0.04 mm diameter) and rounded in shape.
(5) In older animals, chondrocytes form nests or isogenous groups within single lacunae.
(6) With age, the staining of the cartilage matrix by alkaline stains used for microscopy decreases.
(6) Relative to young chondrocytes, older chondrocytes have more stored glycogen.
Unfortunately these things are invisible to the butcher, and seldom examined even by scientists, otherwise they would be very useful in judging a carcass.
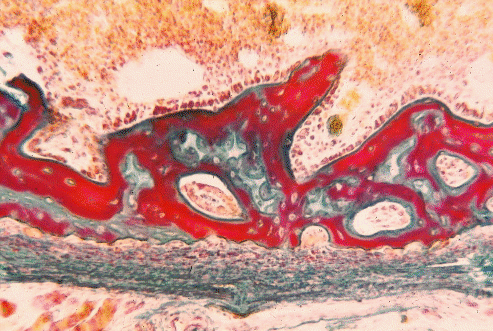 Section of very young bone, with marrow cavity towards the top, and calcified tissue stained red.
Section of very young bone, with marrow cavity towards the top, and calcified tissue stained red.
Oxygen, nutrients and waste products may travel to and from the chondrocytes in cartilage by diffusion through the surrounding matrix but, when the matrix becomes ossified by the deposition of submicroscopic hydroxyapatite crystals, diffusion is greatly reduced. In bone, osteocytes can only survive if they develop root-like cytoplasmic extensions radiating from the lacunae to regions where exchange by diffusion can take place. These cytoplasmic extensions run through fine tubes or canaliculi in the ossified matrix, but are limited in length. Consequently, large numbers of blood vessels permeate the matrix of bone. Most of these blood vessels run longitudinally through the bone in large haversian canals surrounded by concentric rings of osteocytes and bone lamellae. Bones are covered by a connective tissue membrane called the periosteum.
The prenatal formation of bone is initiated by either of two basic mechanisms, (1) intramembranous ossification or (2) endochondral ossification. Intramembramous ossification is typical of the bones that form the vault of the skull, and it occurs when sheets of connective tissue produce osteoblasts which then initiate centers of ossification. Endochondral ossification is more common, and is the process by which cartilagenous models become ossified to form the bones of a commercial meat carcass.
The internal structure of carcass bones becomes visible when they are split longitudinally on a band saw. The shaft of a bone is called the diaphysis, while the knob at each end is called the epiphysis. Between the diaphysis and each epiphysis is a cartilagenous growth plate called the epiphyseal plate. In a young animal, the chondrocytes of the epiphyseal plate are constantly dividing to form new matrix. However, on each face of the plate, cartilage is being continuously resorbed and replaced by bone so that the thickness of the epiphyseal plate tends to remain constant in growing animals. This process allows a bone to grow longitudinally without disrupting the articular surface on the epiphysis. The rate of the longitudinal growth of bones is the product of two factors; (1) the rate of production of new cells, and (2) the size that cells reach before they degenerate at the point of ossification. The strength and thickness of epiphyseal plates is modified by sex hormones. At puberty, chondrocyte growth slows down and fails to keep pace with ossification on the surface of the epiphyseal plate. Thus, epiphyseal plates are lost in mature animals, and the epiphyses become firmly ossified to their diaphyses. However, the factors that regulate the closure of the epiphyseal plate and, hence, the frame size of the animal, are poorly understood at present. Although regulation is likely to be an interaction between animal age and circulating hormones, there are no obvious hormonal changes when the plate closes. If whethers are implanted with the female horme estradiol, the ossification of growth plates is accelerated. Bone growth in mature animals is restricted to the girth or thickness of the bone, and it occurs by the recruitment of periosteal cells to become osteoblasts.
Bones are the main storage site for calcium in the body, and calcium ions are absolutely vital to the normal functioning of body cells. So when calcium is in short supply, because a cow is suddenly producing lots of milk or a hen is laying eggs like crazy, calcium is released from the bones into the blood. Milk fever may occur in a cow if the skeleton cannot keep pace with the demand.
Carcasses from young animals have a relatively high bone content because the skeleton is well developed at birth. As an animal grows to market weight, its proportion of bone decreases on a relative basis, because of the growth of muscle and fat. The long-term control of bone growth is superimposed on the short-term regulation of bone metabolism that occurs in response to changes in blood calcium levels, or to remodelling in response to local functional demands.
A number of hormones exert secondary effects on skeletal development. Thyroxine, insulin, growth hormone and gonadal hormones tend to be anabolic, building up the bone mass. Estrogens may inhibit resorption of bone. Adrenal corticosteroids stimulate resorption of bone and inhibit the formation of new bone. In cattle, castration delays the completion of growth in epiphyseal plates. This is most noticeable in the distal bones of the limbs and enables the continued longitudinal growth of the legs. In the vertebral column, however, castration reduces bone growth. The reduction is centered on the first thoracic vertebra. Removal of the ovaries from heifers also causes an increase in the longitudinal growth of distal bones. Growth factors may mediate or augment the activity of the hormones controlling bone growth. Both rapidly growing and adult bones may contain growth factors such as transforming growth factor beta, beta-2 microglobulin, and insulinlike growth factor I. In adults, these factors may be involved in skeletal remodeling.
There is a lot of uncertainty about the local control of bone growth, something that enables bones to develop struts of strong bone where they are most needed. One hypothesis is that loads frequently placed on a region of bone cause the conversion of mechanical energy to electrical energy by a piezoelectric effect (the same effect used in the crystal oscillator of an electric wrist-watch). In a frequently loaded and negatively charged region of bone, growth is stimulated. Experimentally, the growth of both bone and cartilage may be modified by the application of pulsed magnetic fields and, in electrical fields, osteoclasts that erode bone migrate towards the positive electrode while osteoblasts that deposit bone migrate towards the negative electrode.
In an unloaded and positively charged region, resorption is stimulated. Differences in the arrangement of the hydroxyapatite crystallites of bone, in lacunar structure, and in the transition from spongy to compact bone have been observed between the bones of wild and domesticated sheep. These differences may have accompanied a reduction in exercise during domestication.
The gross anatomy of muscles and skeletal units are closely matched, and mutual or interacting control systems probably exist. Because most farm animals are slaughtered in a fairly immature condition, the relationship between muscles and the bony processes that they pull on may not be immediately obvious. But knobs and wrinkles on bone surfaces become more conspicuous with age, and they are readily seen in the carcasses of old bulls. One possible relationship between muscle activity and bone growth may be that isometric contraction (like in a weight lifter), by stopping or slowing the venous blood flow, may stimulate bone growth. Alternatively, by pulling on the periosteum, the effect of muscle activity may be mediated by connective tissue. The importance of local factors is seen in bone transplants, where growth of the transplant almost immediately becomes regulated by the new local conditions.
Breeds of cattle with a large mature size usually produce lean meat at a faster rate than early maturing traditional beef breeds with a relatively small adult size. Differences in adult size are produced by differences in skeletal growth, and relationships between the quantitative anatomy of individual bones and meat production traits in beef cattle have been identified. Relationships between skeletal and muscular development may involve meat quality because large-framed animals produce leaner meat during their production life span. Large-framed breeds mature late and have a later cessation of linear skeletal growth at their epiphyseal plates. The time of maturation is related to the distribution and amount of adipose tissue in the carcass, particularly marbling fat. Differential bone growth between large and small breeds of cattle is usually established prior to a slaughter weight of 500 kg in males. But the emphasis that is placed on animal height by many beef breeders may be misplaced. Comparing present day cattle with those born 20 years ago, faster growing modern animals may be longer in the body, but not necessarily taller than their predecessors.
Pelvic dimensions in cows of different breeds are related to the incidence of difficult calving or dystocia. Dystocia may be particularly serious when a homozygous double-muscled calf is born. Double-muscling in the calf is caused by increase in the number of muscle fibres so that the shape of the calf is very bulky. The dam, which may be either a heterozygous carrier or completely double muscled, may also exhibit some reduced bone development.
Although one might expect the proportion of bone in a carcass to affect specific gravity measurements, this may not be evident in practice. Growth promotants usually have little or no effect on skeletal development, but environmental factors do affect bone development, since certain confinement conditions cause lameness involving skeletal joints.
In the early 1950s, attempts were made to use measurements of isolated carcass bones such as the cannon bone to predict the muscle to bone ratios of carcasses. Although the method worked satisfactorily when applied to a wide range of dissimilar carcasses, it was of little practical value when applied to more uniform commercial carcasses. Muscle to bone ratios improve as animals grow older or fatter, since longitudinal bone growth slows down in older animals and muscles start to accumulate appreciable amounts of intramuscular fat. Animal age is the dominant factor that determines muscle to bone ratios. However, when adjustments are made for animal age and carcass weight, considerable unexplained variation still is found in muscle to bone ratios.
An emphasis on larger, leaner breeds of cattle has obscured the fact that, some years ago, the trend was in the opposite direction. The desire to produce small compact animals with bulging muscles favored the survival of dwarf animals with impaired longitudinal bone growth. Although mildly affected animals looked very muscular, severely affected animals became increasingly common and were poorly suited for beef production. Dwarfism from impaired longitudinal growth of bones is a recessive trait that affects males more strongly than females.
The meat fragments remaining on bones after they have been boned out may be retrieved by grinding the bones and meat fragments, and then subjecting them to great pressure behind a metal screen or sieve. Residual meat fragments are pushed through the screen and are collected as mechanically deboned meat. Mechanical deboning enables the recovery of meat from bones such as vertebrae that are difficult to clean manually. However, very small bone particles and fat also are extruded through the screen, together with the meat particles. With a hole size of 0.46 mm in the screen, bone particles from beef neck muscles ranged in size from 0.08 to 0.11 mm. Bone particles of this size are dissolved in the acidic conditions found in the adult human stomach. Residual calcium content normally is maintained at relatively low levels in mechanically deboned meat. Mechanically deboned meat is generally used as a supplement in processed meat products.