19
Fatty acid metabolism
19.1 Introduction
Adipose cells store energy as triglyceride. A molecule of triglyceride is
composed of three fatty acids on a glycerol backbone like a letter
E. Sometimes the fatty acids pass through pigs and poultry from
their feed without much change, and the resulting fat can develop a
terrible aroma. There is no point in producing meat we cannot
sell. Thus, animal science students need to understand how this
happens.
- Adipose
tissue depots are
primarily for energy storage
or
for mechanical and thermal insulation, but
they are very active metabolically.
- Anabolic
(building-up) processes are very active when an animal is depositing
fat, while catabolic
(breaking down) processes are very active when an animal is living off
its energy reserves,
either during prolonged starvation or between periods of feeding.
- For
example, the
metabolism of adipose cells is altered in cows after calving, due to
the
mobilization of adipose reserves for milk production: lipogenesis (building-up lipid) is
reduced
and lipolysis (breaking-down
lipid) is increased.
- Also, metabolic water may be
produced by oxidation of fat
when animals are deprived of water (remember the hump on the camel is
fat - not water!). Thus, the
depth of backfat and the total weight of subcutaneous fat may be
reduced in
pigs on a restricted water intake.
IN THE DIAGRAMS - NOTHING IS TO
SCALE. ORGANS ARE ALWAYS BIGGER THEN CELLS, AND CELLS ARE
ALWAYS BIGGER THAN MOLECULES - BUT
NOT IN THESE DIAGRAMS!
19.2
Structure of triglyceride

The long‑chain fatty
acids found in
animal triglycerides vary in length, as shown by
the values of "n" in the general formula for fatty acids. The value
for "n" in animal fat usually ranges from 5 to 20. These fatty acids
are insoluble in water, like the triglycerides formed from them.
Adjacent
carbon atoms along a chain may be linked together with an unsaturated double
bond. Unsaturated bonds produce a bend in the chain and this lowers the melting
point. Unless complicated by the presence of unsaturated bonds, melting points
are proportional to chain length.
General structure CH3‑(CH2)n‑COOH
Names for values of "n"
- not counting the
carbons at the ends of the chain!
2 ‑
butyric
4 ‑
caproic
6 ‑
caprylic
8 ‑
capric
10 ‑ lauric
12 ‑ myristic
14 ‑ palmitic
16 ‑ stearic
18 ‑ arachidic
20 ‑ behenic
The degree of saturation of
carcass fat is affected by a number of factors.
- Within a
breed, steers may have
more saturated fat than heifers but differences may also
exist between breeds, as in comparisons of Japanese Wagyu cattle with
Angus (this is important if we are trying to produce beef which will
appeal to Japanese consumers!).
- About 12
months after birth, cattle normally start to
fatten rapidly, and progressively more unsaturated fatty acids are
deposited.
- Genetically
fat pigs have a greater degree of saturation
of their fatty acids than normal pigs.
- Most
plant oils contain a high
proportion of unsaturated fatty acids with low melting points, but
animals have
a high proportion of saturated fatty acids and their fat is solid or
semi‑solid
at room temperature.
- Thus, when a beef carcass is cooled
from body temperature
down to nearly 0oC after slaughter, subcutaneous fat
passes from a
liquid to a solid state.
- In beef
carcasses, a greater proportion of fatty acids
may be saturated in the summer than in the winter. Seasonal
changes of this type are quite marked in wild sheep, where they may
help the
animal to adapt to climatic changes.
- Fat
tends to be soft and
yellow in ram carcasses relative to wethers, and fat also tends to be
soft and
yellow in lambs fed a high‑energy diet relative to those on a
low‑energy diet.
- In
general, triglycerides
located subcutaneously where they may be relatively cool in the live
animal
have a low melting point. Conversely, perirenal or suet fat in the beef
carcass
is brittle at room temperature since it comes from a warm place in the
body and
has a high melting point.
- In
animal fat, most fatty
acids contain an even number
of carbon atoms, and unsaturated bonds are either
absent or few in number. In meat, the most common unsaturated fatty
acids are oleic, linoleic and linolenic acids with 18 carbons and
with 1, 2, and 3 double
bonds, respectively.
19.3
Digestion and deposition of triglyceride
The
integration of adipose
metabolism with that of the rest of the body is VERY complicated.
THIS IS THE SESAME STREET VERSION.
Dietary
intake
In the case of a pig with triglyceride in its diet, the
triglyceride reaches the small
intestine and
is hydrolyzed in the presence of pancreatic lipase.
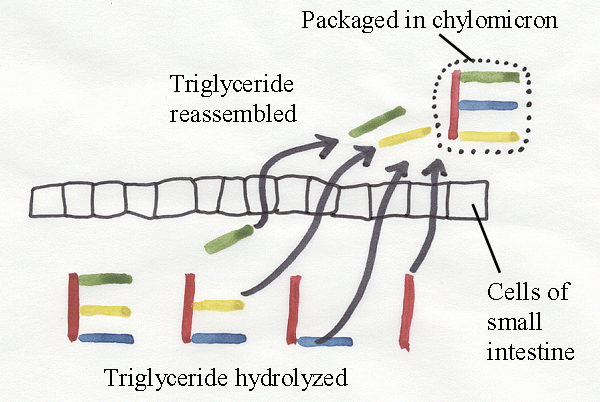
Colipase, a
protein secreted by the pancreas,
binds to the surface of fat droplets
in the
presence of bile salts from the liver and facilitates the attachment of
lipase. The partially or completely separated fatty
acids may
pursue a number of different pathways, one of which involves absorption
by an
intestinal cell and reassembly into a new triglyceride inside the cell.
Large
numbers of reassembled triglyceride molecules may be wrapped in protein
and
phospholipid to form a small globule about one micron in diameter
called a
chylomicron.
The triglycerides
contained in the chylomicra then pass to
the
lacteals. Lacteals are
blind‑ending lymphatic
capillaries in the
axes of the intestinal villi.
The lymphatic system conducts chylomicra
to the
venous system where, after
passing into the general circulation, they
are made
available to adipose cells.
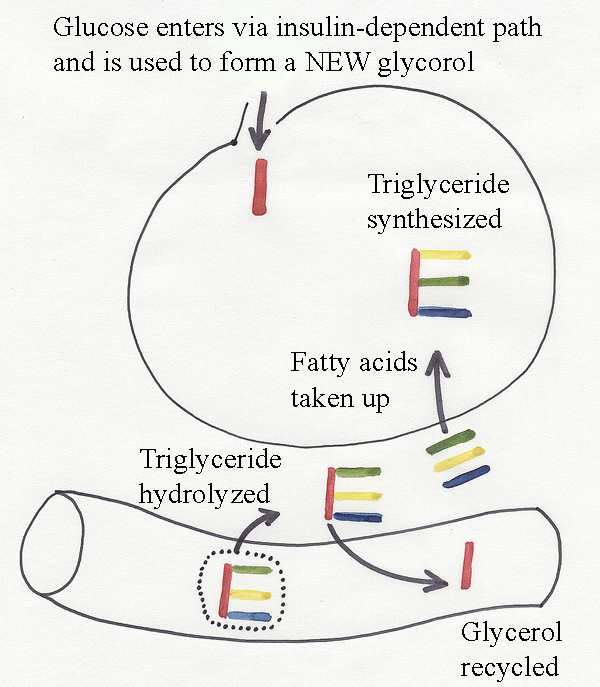
However, chylomicra do
not
enter
directly into adipose cells. Their triglyceride is first hydrolyzed to
fatty
acids and glycerol by lipoprotein
lipase (clearing factor lipase)
located on
the surfaces of adipose cells or capillary cells. The
transfer
of the fatty acids to the adipose cell may be facilitated by surface
proteoglycans trapping
lipoproteins together with lipases, and by the
continuity
of intercellular connections with intracellular channels. Yet again,
triglycerides now are reassembled, this time
within the
adipose cell. Lipoprotein lipase activity in adipose
tissue is
linearly related to cell size.
Most
of the glycerol released
by lipoprotein lipase outside the adipose cell is recycled to the blood
stream. Inside the adipose cell, a new carbon backbone for the
triglyceride is formed from glycerophosphate derived from
glucose. The entry of glucose into a cell is facilitated by
insulin. In
porcine adipose tissue the enhancement of carbohydrate metabolism by
insulin is
antagonized by porcine growth hormone (STH). This
may
explain the leaner carcasses and greater lean growth of pigs treated
with
growth hormone.
- By a
simple route, such as
that outlined above, ingested fatty acids may be deposited in an
unaltered form
in their new "host".
- Thus, in
pigs, the degree of saturation of
carcass fat is influenced by the nature of the fatty acids
ingested.
- This may
affect meat quality because fat with a low melting point
appears
greasy.
- Fat with a relatively large number of
unsaturated bonds also is
more
likely to undergo auto‑oxidation with the formation of unpleasant odors.
- Commercial
problems often arise when highly
unsaturated
fats
such as linseed oil or fish oil are fed to monogastric animals, since
these
oils may cause unpleasant odors in the meat.
- Unpleasant
odors may not
become
apparent until the meat has been cooked and its unsaturated fatty acids
have
been oxidized.
- Such
problems are rare in ruminants
since
unsaturated fats are reduced in the rumen. However, prior treatment of
the feed
with formaldehyde inhibits this reduction and enables unsaturated fatty
acids
to pass through the digestive system relatively unchanged.
- In
cattle and sheep, dietary lipids are extensively modified in the
rumen.
Esterified fatty acids are release by hydrolysis, the residual glycerol
and
galactose are fermented, and unsaturated fatty acids are hydrogenated.
- Large
amounts of stearic acid are
formed from oleic, linoleic
and
linolenic acids. These, together with other fatty acids derived
from
the
microflora, are absorbed in the intestine.
- In pigs,
a high linoleic intake in
the diet is
notorious for softening the subcutaneous fat of the carcass
and
causing problems for meat cutters.
- There is
also the possibility
of using the direct dietary incorporation of fatty acids to some
advantage, as
in the case of 20-carbon omega-3 fatty acids from marine sources. These have acquired a reputation for being a
desirable component of the human diet. The
basic problem is the inclusion of marine oils into
animal feeds
causes off-flavors in the product.
19.4 Lipogenesis
An alternative carbon input is
available to adipose cells from very-low-density
lipoproteins (VLDL)
from the
liver. In VLDL, a central core of
triglyceride is
packaged by a layer of protein,
phospholipid and cholesterol. The
leaching of
triglyceride by lipoprotein lipase
near to adipose cells leads
to a reorganization of VLDL components, and a low-density lipoprotein
(LDL) is
produced. The cholesterol remaining in LDL is released into the
blood stream
by cells of the reticuloendothelial
system. In the
liver,
cholesterol, triglyceride and circulating high-density lipoproteins
(HDL) are
assembled into new VLDL.
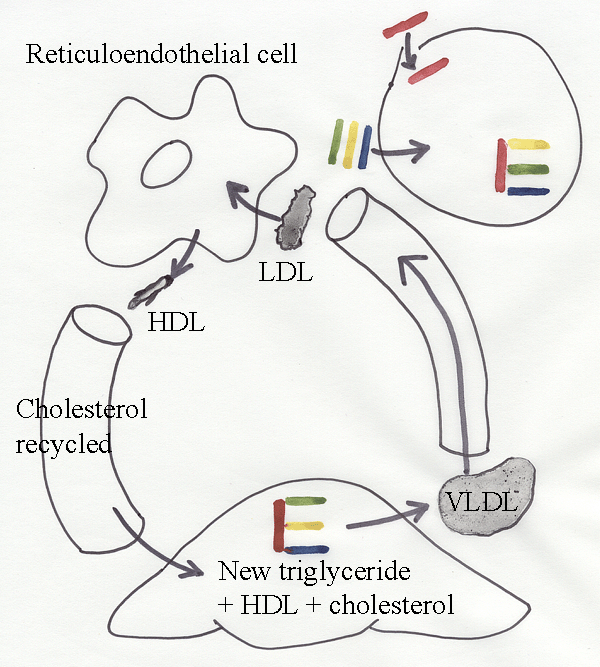
Meat
animals may form new fat
by lipogenesis, as shown in
the classical experiments of Lawes and
Gilbert in Victorian England. In pigs, most of the new triglyceride is
synthesized from
glucose, and the fatty acids produced are dominated by stearic
acid
and its desaturated form, oleic
acid. In ruminants, acetate
is the main
substrate, and stearic acid is
the dominant fatty acid.
In poultry,
lipogenesis
occurs in the liver rather than in adipose cells, and
triglyceride
reaches the
adipose cells as VLDL in the blood. Oleic and palmitic acids are
dominant when
the dietary intake of triglyceride is low.
Further information
Structure
and Development of Meat Animals and Poultry. Pages 116-120.